
Una nutrición correcta favorece al disfrute del más alto nivel posible de salud física y mental de niñas y niños, la malnutrición por lo tanto afecta al desarrollo de los niños y niñas de contexto escolar en todas sus facetas de crecimiento y desarrollo.
Hay diversos factores que pueden tener un rol en el incremento de peso y este proceso metabólico llevar a la obesidad y por consecuencia sufrir enfermedades metabólicas crónicas, pero también puede tener un papel importante en la disminución de su crecimiento y desarrollo a causa de una deficiencia alimentaria u otros factores que contribuyan a la mal nutrición de los mismos en etapas de evolución de crecimiento.
En el artículo Epidemiología de las enfermedades metabólicas resultantes de la malnutrición: El caso de México, del Dr. Carlos A. Aguilar Salinas plasmo el proceso en la epidemiologia de las enfermedades metabólicas, el incremento de la población en México, la modificación de las conductas alimentarias de la población Mexicana, así como la epidemiologia de la obesidad de los mismos.
Los datos que aquí integra demuestran que la epidemiología de las enfermedades metabólicas cambió en México en un periodo de cuatro décadas (1950-1990) siendo más notables las enfermedades crónicas degenerativas por el aumento en los índices de masa corporal.
La dinámica de la población modificó las cadenas alimentarias, la actividad física de la población y la estructura de las familias. Como resultado, la alimentación sufrió modificaciones profundas, caracterizadas por aumento en el aporte de calorías, grasas saturadas y azúcares simples provocando en la población mexicana un aumento de peso en niños, adolescentes y adultos, lo que por consecuencia ha acarreado enfermedades crónicas degenerativas y muertes en personas de edades más tempranas.

¿Cuáles son las cifras en el aumento de peso?
La obesidad afecta a todos los grupos etarios. El porcentaje de niños entre 5 y 11 años con obesidad creció de 14.6% en 2006 a 17.5% en 2018. Al considerar a los niños en riesgo de obesidad (categoría equivalente al sobrepeso en adultos), el 35.6% tiene un peso mayor al saludable. En los adolescentes (12 a 19 años), el porcentaje con un peso mayor al saludable creció de 34.9 en 2012 (21.6% sobrepeso y 13.3% obesidad) a 38.4% en 2018 (23.8% sobrepeso y 14.6% obesidad). Al igual que en los adultos, la proporción de los casos con un peso mayor al saludable es mayor en las mujeres (sobrepeso 27%, obesidad 14.1%, total 41.1%) que en los hombres (15.1%). Todo esto llevo al incremento de enfermedades y aumento en el costo de hospitalizaciones y muertes a edades tempranas.
¿Qué estrategias se implementaron para prevenir y ayudar a reducir estas cifras?
A finales de 2015, miembros del Área de Investigación en Políticas y Programas de Nutrición del Instituto Nacional de Salud Pública (INSP) dieron a conocer resultados preliminares de estudios que abordan importantes aspectos de la nutrición poblacional desde la óptica de los programas y políticas públicas.
Los resultados de las investigaciones se presentaron a través de carteles y exposiciones orales durante la reunión anual Obesity Week 2015 en la ciudad de Los Ángeles. Asimismo, durante el evento el director de Investigación en Políticas de Nutrición del INSP, Dr. Simón Barquera, impartió dos conferencias magistrales durante las sesiones Towards global monitoring of food environments and policies: the INFORMAS initiative y TOS Latin American Affairs Section Meeting.
Entre los temas investigados se incluyen las estrategias de publicidad de alimentos y bebidas dirigidas a la población infantil en diversos canales de comunicación y su frecuencia; la calidad nutrimental de estos productos (contenido de sodio, azúcar y grasa); un programa de reducción de peso utilizando la metodología de Silicon Valley (CARTEL: Evolutionary Designs: Using Silicon Valley Methodologies to Develop an Intervention for Weight-Loss Under Economic Constraints (1),de Victor Villalobos e Ismael Campos. Presentó la investigación realizada para el desarrollo e implementación de un programa de reducción de peso aplicable en comunidades de bajo ingreso en zonas rurales. El programa desarrollado se implementó por primera vez en mayo del 2015, incluyendo componentes como asesoría nutricional y de salud, herramientas de auto monitoreo, búsqueda activa de apoyo social, creación de objetivos, métodos de compromiso y asistencia remota. Ganador del 4th Annual Latin American Affairs Poster Award.).
Y resultados de la evaluación de implementación del Programa de Acción en el Contexto Escolar (Cartel. Evaluation of Two Implementation Strategies of the School Context Action Program (PACE, According to its Initials in Spanish) in Mexico (2), de Lizbeth Tolentino-Mayo, Lucía Hernández, Enrique Cifuentes, Lucie Lévesque y Simón Barquera. Reportó resultados de la evaluación de la implementación del Programa de Acción en el Contexto Escolar (PACE). Los resultados refieren a una comparación entre dos periodos de implementación en 20 escuelas del Distrito Federal. Entre estos periodos, se observó una reducción en la cantidad de sodio en los alimentos preparados que se ofrecían en el recreo, de 248.7 a 204.4 mg/porción. Asimismo, se identificó una disminución en la cantidad de calorías por porción en las botanas saladas pasando de 116.7 a 43.6 kcal/porción, aunado a una constante disponibilidad de frutas y verduras durante la semana en el 80% de las escuelas. Ganador del 4th Annual Latin American Affairs Poster Award).
Este último arrojo datos interesantes que deja en visto que la implementación de estrategias de publicidad Nutricionales para prevención de la mal nutrición son muy eficientes, especialmente si son dirigidos en los grupos con mayor aprendizaje y vulnerabilidad, que es la población en el contexto escolar.
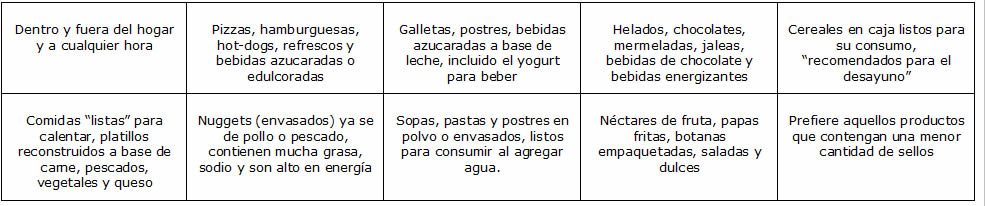
Pero también podemos percibir otros panoramas de mal nutrición que no solo son por obesidad sino también por deficiencia calórica, vitamínica y proteica que retarda el crecimiento de niños y niñas generando también enfermedades crónicas degenerativas como es en el caso de las zonas más vulnerables y marginadas de nuestro país en donde el acceso a los alimentos es limitado y difícil de obtener. Sin embargo hay factores externos que influyen a la mal nutrición como es el caso de la contaminación de agua, la falta de higiene, el exceso de químicos para su producción de alimentos en el campo por mencionar algunos otros.
La malnutrición es un problema que genera muertes y en el largo plazo afecta también a quienes sobreviven como es el caso de los niños con insuficiencia renal de la Ribera de Chapala y otras zonas cercanas que día a día están perdiendo la batalla contra esta enfermedad.
En Jalisco en el año 2019 la Secretaria de Educación Jalisco implementó un programa llamado Recrea Escuela para la vida a manera de programa piloto, donde se tomaron en cuenta a 50 escuelas primarias públicas del estado de Jalisco en regiones altos, sur, Ciénega, ribera y zona metropolitana de Guadalajara, el instituto de Nutrición del Dr. Salvador Zubiran realizo el diagnostico basal de inicio, lamentablemente no fue concluido este programa a causa de los estragos de la pandemia que por consecuencia dejo un bajo impacto en los resultados de estas escuelas pues tuvieron que cerrar y hasta la actualidad no han dado indicios de que se tenga la intención de retomar este estudio, sin embargo se puede afirmar que lo poco que se llevó a cabo fue de interés para los padres de familia y los niños de estas escuelas primarias. Se notó un comportamiento alimentario modificado en los niños pero a nivel de índice de masa corporal no hubo estudio que revelaran resultados de dicha modificación. La intención de estos programas es corregir y erradicar esta enfermedad que les aqueja, para disminuir la gran tasa de muerte que surgió en niños y jóvenes de esta región.
Y así como estos programas se sabe que hay más similares con las mismas estrategias que no se plasman en este escrito pero se deja la invitación para seguir investigando y poniendo en marcha toda estrategia que pueda apoyar a nuestro país a mejorar el estado de salud de cada niño. Por consecuente se sabe que la toma de dichas estrategias demuestra ser herramientas óptimas para apoyar y modificar comportamientos que provocan aumento de peso y por consecuencia enfermedades degenerativas, pero también son de ayuda para prevenir alteraciones de mal nutrición en la población infantil mexicana.
PARA LEER MÁS, FUENTES:
https://www.insp.mx/epppo/blog/3978-nuevas-investigaciones-nutricion.html
Epidemiología de las enfermedades metabólicas resultantes de la malnutrición: El caso de México
https://proyectoeducativo.jalisco.gob.mx/escuela-para-la-vida








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}